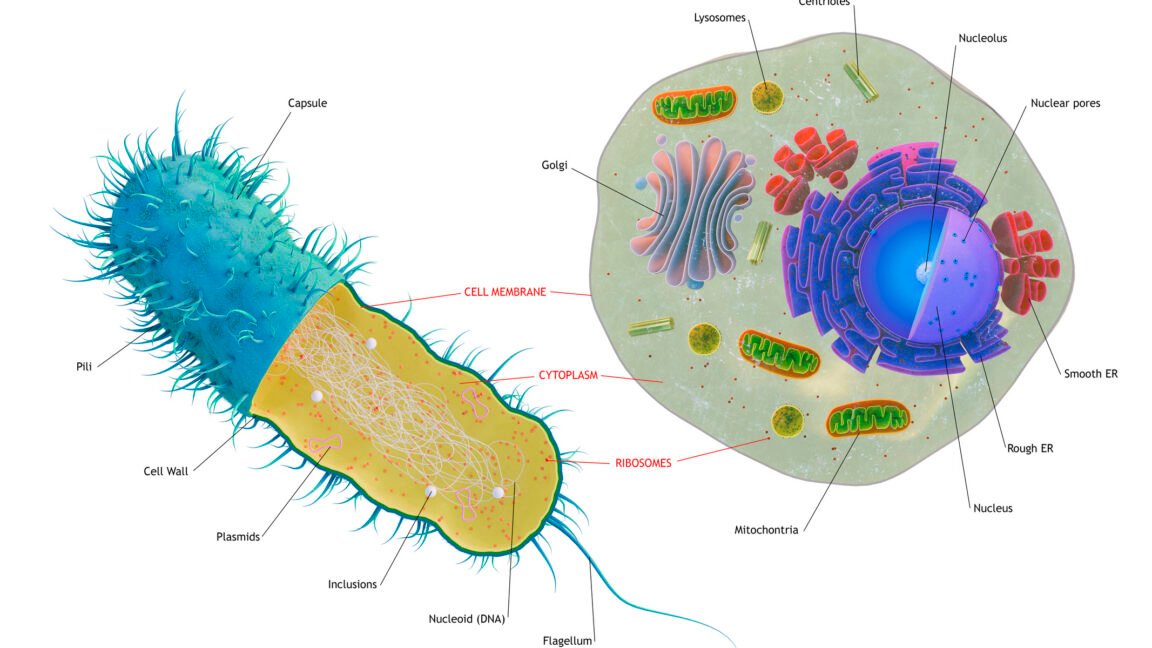
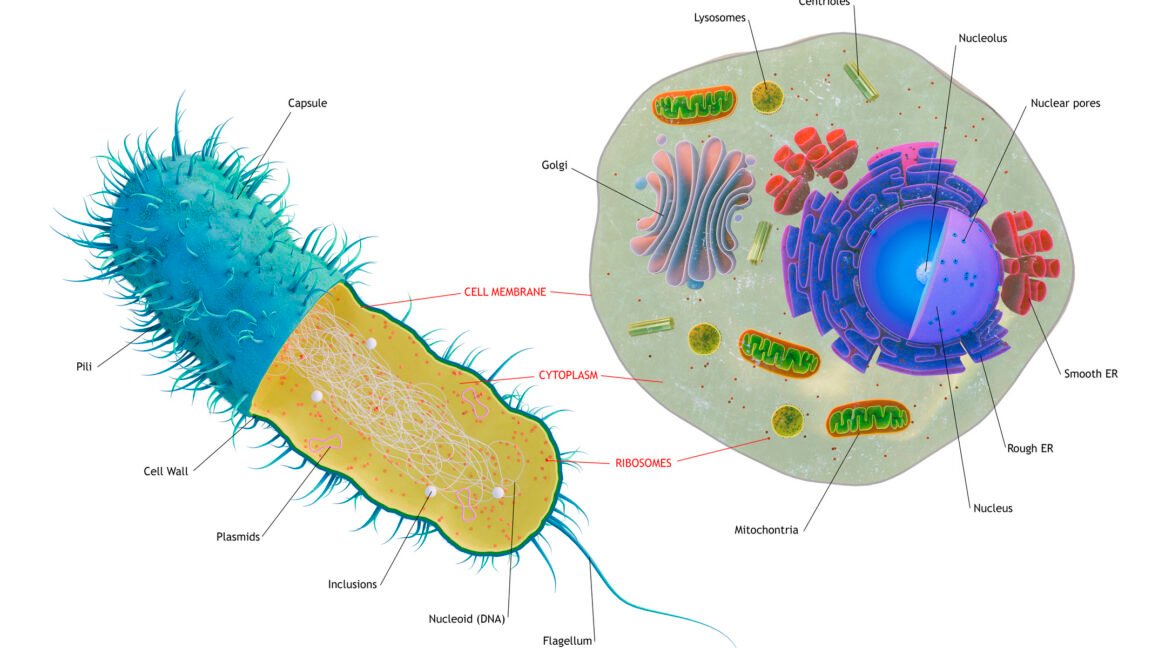
ان خلیات میں پہلے سے ہی پیچیدہ اندرونی حصے ہوتے ہیں، جس میں اندرونی پروٹین ٹریک ویز موٹر پروٹینوں سے گزرتے ہیں جو سیل کے اندر کارگو کو منتقل کرتے ہیں۔ خلیوں کے اندر پروٹین کو ہضم کرنے کے لیے ڈھانچے (لائسوزوم اور پیروکسوم) تھے، اور یوکرائیوٹک میٹابولزم، ڈی این اے کی نقل، اور آر این اے کی پیداوار کی تمام بنیادی باتیں۔ ایک بڑی خصوصیت جو غائب تھی وہ جینوں کے سیٹ تھے جو اس بات کا تعین کرنے کے لیے استعمال کیے جاتے تھے کہ ایک خلیے کو کب تقسیم ہونا چاہیے اور ان واقعات کا انتظام کرنا چاہیے جو اس کے لیے ہونے کی ضرورت ہیں۔ یہ تجویز کر سکتا ہے کہ سیل ڈویژن صرف میٹابولک خدشات کی وجہ سے محدود ہے۔
یہ کیسے ہوا؟
تقریباً ایک تہائی جین گروپ یوکرائٹس سے الگ نظر آتے ہیں اور دوسری ریاستوں میں ان کے مساوی نہیں ہیں۔ ان میں سے کچھ اس نسب میں موجود ہو سکتے ہیں جس نے یوکرائٹس کے آخری مشترکہ اجداد کو پیدا کیا تھا، اور کچھ اس سے پہلے پیدا ہوئے ہوں گے کہ یوکرائٹس نے واقعی متنوع ہونا شروع کر دیا ہو اور شاخیں نکلیں۔
جیسا کہ توقع کی گئی ہے، بہت سے دوسرے جینز یا تو Asgard archaea یا Alphaproteobacteria سے آئے ہیں، جو ہماری اصلیت کے بڑے تصویری ماڈل کے مطابق ہیں۔ لیکن محققین نے دو دیگر بیکٹیریل گروپس سے بھی تقریباً مساوی شراکت پائی: پلانکٹومیسیٹوٹا اور مائکسکوکوٹا۔ (اس میں شامل تمام بیکٹیریل گروپ متنوع اور نسبتاً عام ہیں، اسگارڈ آثار قدیمہ کے بالکل برعکس)
مختلف بیکٹیریل گروپس کی ایک رینج کی طرف سے چھوٹی چھوٹی شراکتیں بھی تھیں۔ لیکن وائرس کے گروپ کی پرجاتیوں میں شامل ہیں۔ وشال وائرس کسی ایک بیکٹیریل گروپ سے زیادہ حصہ ڈالا۔
محققین نے اس وقت کا بھی اندازہ لگایا کہ جب جین کے گروپ متعارف کرائے گئے تھے۔ Asgard archaea سب سے قدیم شراکت کی نمائندگی کرتا ہے، جیسا کہ توقع کی جائے گی۔ لیکن ایک بیکٹیریل نسب تھا جس نے مائٹوکونڈریا کے موجود ہونے سے پہلے بہت سارے جین متعارف کرائے تھے اور ایک دوسرا گروپ تھا جس نے بعد میں اہم کردار ادا کیا۔ یہ سمجھ میں آتا ہے اگر eukaryotes ایک مائکروبیل چٹائی کے اندر تیار ہوئے ہیں، جہاں بہت سی انواع طویل عرصے تک قریب رہتی ہیں اور بعض میٹابولائٹس کے لیے ایک دوسرے پر انحصار کر سکتی ہیں۔


